Биология - Экосистема - Механизмы функционирования экосистемы
09 февраля 2011Оглавление:
1. Экосистема
2. История термина
3. Понятие экосистемы
4. Строение экосистемы
5. Механизмы функционирования экосистемы
6. Пространственные границы экосистемы
7. Временные границы экосистемы
8. Ранги экосистем
9. Искусственные экосистемы
10. Понятия, сходные с понятием экосистемы, в смежных науках
Устойчивость экосистем

Экосистема может быть описана комплексной схемой прямых и обратных связей, поддерживающих гомеостаз системы в некоторых пределах параметров окружающей среды. Таким образом, в некоторых пределах экосистема способна при внешних воздействиях поддерживать свою структуру и функции относительно неизменными. Обычно выделяют два типа гомеостаза: резистентный — способность экосистем сохранять структуру и функции при негативном внешнем воздействии и упругий — способность экосистемы восстанавливать структуру и функции при утрате части компонентов экосистемы. В англоязычной литературе используются сходные понятия: локальная устойчивость — англ. local stability и общая устойчивость — англ. global stability.

Иногда выделяют третий аспект устойчивости — устойчивость экосистемы по отношению к изменениям характеристик среды и изменению своих внутренних характеристик. В случае, если экосистема устойчиво функционирует в широком диапазоне параметров окружающей среды и/или в экосистеме присутствует большое число взаимозаменяемых видов, такое сообщество называют динамически прочным. В обратном случае, когда экосистема может существовать в весьма ограниченном наборе параметров окружающей среды, и/или большинство видов незаменимы в своих функциях, такое сообщество называется динамически хрупким. Необходимо отметить, что данная характеристика в общем случае не зависит от числа видов и сложности сообществ. Классическим примером может служить Большой Барьерный риф у берегов Австралии, являющийся одной из «горячих точек» биоразнообразия в мире — симбиотические водоросли кораллов, динофлагелляты, весьма чувствительны к температуре. Отклонение от оптимума буквально на пару градусов ведёт к гибели водорослей, а до 50-60 % питательных веществ полипы получают от фотосинтеза своих мутуалистов.
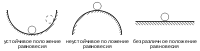
У экосистем существует множество состояний, в которых она находится в динамическом равновесии; в случае выведения из него внешними силами, экосистема совершенно необязательно вернётся в изначальное состояние, зачастую её привлечёт ближайшее равновесное состояние, хотя оно может быть очень близким к первоначальному.
Биоразнообразие и устойчивость в экосистемах

Обычно устойчивость связывали и связывают с биоразнообразием видов в экосистеме, то есть, чем выше биоразнообразие, чем сложнее организация сообществ, чем сложнее пищевые сети, тем выше устойчивость экосистем. Но уже 40 и более лет назад на данный вопрос существовали различные точки зрения, и на данный момент наиболее распространено мнение, что как локальная, так и общая устойчивость экосистемы зависят от значительно большего набора факторов, чем просто сложность сообществ и биоразнообразие. Так, на данный момент с повышением биоразнообразия обычно связывают повышение сложности, силы связей между компонентами экосистемы, стабильность потоков вещества и энергии между компонентами.

Важность биоразнообразия состоит в том, что оно позволяет формировать множество сообществ, различных по структуре, форме, функциям, и обеспечивает устойчивую возможность их формирования. Чем выше биоразнообразие, тем большее число сообществ может существовать, тем большее число разнообразных реакций может осуществляться, обеспечивая существование биосферы в целом.
Сложность и устойчивость экосистем
На данный момент не существует удовлетворительного определения и модели описывающей сложность систем и экосистем в частности. Существует два широко распространённых определения сложности: колмогоровская сложность — слишком специализированное для применения к экосистемам. И более абстрактное, но тоже неудовлетворительное определение сложности, данное И. Пригожиным в работе «Время, хаос квант»: Сложные системы — не допускающие грубого или операционального описания в терминах детерминистских причинностей. В других своих трудах И. Пригожин писал, что не готов дать строгого определения сложности, поскольку сложное — это нечто, что на данный момент не может быть корректно определено.
Параметры сложности и их влияние на устойчивость
В качестве параметров сложности экосистем традиционно подразумевались общее число видов, большое число взаимодействий между видами, сила взаимодействий между видами и популяциями и различные сочетания этих характеристик. При дальнейшем развитии этих представлений появилось утверждение, что чем больше путей переноса и преобразования энергии в экосистеме, тем она устойчивей при различных видах нарушений.
Однако на данный момент показано, что данные представления не могут охарактеризовать устойчивость экосистем. Существует множество примеров как весьма устойчивых монокультурных сообществ, так и слабоустойчивых сообществ с высоким биоразнообразием. Так как существуют природные, почти повсеместные, подтверждения отклонений от принятых воззрений, в 70-80-х годах XX столетия происходили исследования по моделированию зависимости устойчивости от сложности экосистем. Эти модели показали, что в случайным образом генерируемой сети взаимодействия в сообществе при удалении бессмысленных цепей локальная устойчивость падает с увеличением сложности. Если продолжить усложнение модели и учесть, что консументы испытывают влияние пищевых ресурсов, а пищевые ресурсы от консументов не зависят, то устойчивость не зависит от сложности, либо также падает с её увеличением. Это справедливо для детритных цепей питания.
При изучении общей устойчивости на модели из 6 видов исследовалось удаление одного из видов. В качестве параметра устойчивости была принята связность. Сообщество считалось устойчивым, если остальные виды оставались локально устойчивыми. Полученные результаты согласовывались с общепринятыми воззрениями о том, что с повышением сложности при выпадении хищников высшего порядка устойчивость сообщества падает, но при выпадении оснований пищевой цепи с повышением сложности устойчивость повышалась.
В случае упругой устойчивости, когда под сложностью также понимается связность, с повышением сложности упругая устойчивость также повышается. То есть, большее разнообразие видов и большая сила связи между ними позволяет сообществам быстрее восстанавливать свою структуру и функции. Данный факт подтверждает общепринятые взгляды на роль биоразнообразия как некоего пула для восстановления полноценной структуры как экосистем, так и более высокоорганизованных структур биосферы, а также самой биосферы в целом. На данный момент общепринятым и фактически неоспариваемым является представление о том, что биосфера эволюционировала в сторону увеличения биоразнообразия, ускорения обращения вещества между компонентами биосферы, и «убыстрения» времени жизни как видов, так и экосистем.
Потоки вещества и энергии в экосистемах
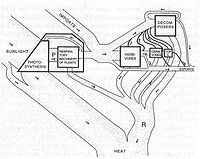
На данный момент научное понимание всех процессов внутри экосистемы далеко от совершенства, и в большей части исследований либо вся экосистема, либо некоторые её части выступают в качестве «чёрного ящика». В то же время, как любая относительно замкнутая система, экосистема характеризуется входящим и выходящим потоком энергии и распределением этих потоков между компонентами экосистем.
Продуктивность экосистем
При анализе продуктивности и потоков вещества и энергии в экосистемах выделяют понятия биомасса и урожай на корню. Под урожаем на корню понимается масса тел всех организмов на единице площади суши или воды, а под биомассой — масса этих же организмов в пересчёте на энергию или в пересчёте на сухое органическое вещество. К биомассе относят тела организмов целиком, включая и витализированные омертвевшие части и не только у растений, к примеру, кора и ксилема,но и ногти и ороговевшие части у животных. Биомасса превращается в некромассу только тогда, когда отмирает часть организма или весь организм. Часто зафиксированные в биомассе вещества являются «мёртвым капиталом», особенно это выражено у растений: вещества ксилеммы могут сотнями лет не поступать в круговорот, служа только опорой растения.
Под первичной продукцией сообщества понимается образование биомассы продуцентами без исключения энергии, затраченной на дыхание за единицу времени на единицу площади.
Первичную продукцию сообщества разделяют на валовую первичную продукцию, то есть всю продукцию фотосинтеза без затрат на дыхание, и чистую первичную продукцию, являющуюся разницей между валовой первичной продукцией и затратами на дыхание. Иногда её ещё называют чистой ассимиляцией или наблюдаемым фотосинтезом).
Чистая продуктивость сообщества — скорость накопления органического вещества, не потребляемого гетеротрофами. Обычно вычисляется за вегетационный период либо за год. Таким образом, это часть продукции, которая не может быть переработана самой экосистемой. В более зрелых экосистемах значение чистой продуктивости сообщества стремится к нулю.
Вторичная продуктивность сообщества — скорость накопления энергии на уровне консументов. Вторичную продукцию не подразделяют на валовую и чистую, так как консументы только потребляют энергию, усвоенную продуцентами, часть её не ассимилируется, часть идёт на дыхание, а остаток идёт в биомассу, поэтому более корректно называть её вторичной ассимиляцией.

Распределение энергии и вещества в экосистеме может быть представлено в виде системы уравнений. Если продукцию продуцентов представить как P1, то продукция консументов первого порядка будет выглядеть следующим образом:
- P2=P1-R2,
где R2 — затраты на дыхание, теплоотдача и неассимилированная энергия. Следующие консументы переработают биомассу консументов первого порядка в соответствии с:
- P3=P2-R3
и так далее, до консументов самого высшего порядка и редуцентов. Таким образом, чем больше в экосистеме потребителей, тем более полно перерабатывается энергия, первоначально зафиксированная продуцентами в пластических веществах. В климаксных сообществах, где разнообразие для данного региона обычно максимально, такая схема переработки энергии позволяет сообществам устойчиво функционировать на протяжении длительного времени.
Энергетические соотношения в экосистемах
- Соотношения B/R и P/R. Первое соотношение показывает необходимое количество энергии, затрачиваемой на поддержание существующей биомассы. В случае, если сообщество находится в критических условиях, данное соотношение уменьшается, так как необходимо затратить больше энергии на поддержание той же биомассы. Обычно в таких ситуациях биомасса также уменьшается. Второе соотношение, величина, характеризует эффективность затрачиваемой энергии на производство биомассы. Такое соотношение можно наблюдать в популяции мелких млекопитающих.

- Соотношения A/I и P/A. Первое называется эффективностью ассимиляции, а второе — эффективностью роста тканей. Эффективность ассимиляции обычно варьирует от 1-4 % для растений и до 20-60 % для животных, для которых всё определяется качеством пищи: травоядные обычно усваивают не более 10-15 % поступившей энергии. Хищники, наоборот, могут ассимилировать до 60-90 %. Эффективность роста тканей в большой степени зависит таже и от коэффициента P/R и изменяется в довольно больших пределах. Поэтому, как и P/R, эффективность роста тканей обычно достигает максимальных значений в популяциях малых организмов, в условиях, где не требуется больших затрат на поддержание гомеостаза.
- Соотношение P/B является важной характеристикой зрелости сообщества. Коэффициент P/B безразмерен и может рассчитываться как продукция за определённый промежуток времени к средней за этот промежуток биомассе, или в конкретный момент времени как продуктивность в этот момент к существующей биомассе. Это соотношение обычно намного больше единицы в молодых сообществах, но с ростом числа видов и приближением к климаксному сообществу этот коэффициент стремится к единице.
Просмотров: 66050
|
|