Биология - Протеинкиназы - Серин/треонин — специфичные протеинкиназы
08 февраля 2011Оглавление:
1. Протеинкиназы
2. Тирозиновые протеинкиназы
3. Серин/треонин — специфичные протеинкиназы
4. Гистидин — специфичные протеинкиназы
Серин-треониновые протеинкиназы фосфоририруют гидроксильную группу в остатках серина или треонина.
Активность этих протеинкиназ регулируется несколькими событиями, а также некоторыми химическими сигналами, в том числе, cAMP, cGMP, диацилглицеролом, Caкальмодулином.
Серин/треониновые протеинкиназы фосфорилируют остатки серина или треонина в консенсусных последовательностях, которые образуют фосфоакцепторный сайт. Эта последовательность остатков аминокислот в молекуле субстрата, позволяет осуществлять контакт каталитической щели протеинкиназы с фосфорилируемой областью. Эта особенность делает киназу специфичной не к какому-либо определенному субстрату, но к специфичному семейству белков с одинаковыми консенсусными последовательностями. В то время, как каталитические домены этих протеинкиназ высококонсервативны, последовательности узнавания отличаются, обуславливая узнавание разных субстратов.
Киназа фосфорилазы была открыта Кребсом в 1959 году и является первым описанным ферментом семейства серин/треониновых протеинкиназ. Киназа фосфорилазы превращает неактивную гликогенфосфорилазу В в активную форму гликогенфосфорилазу A, последняя отщепляет от гликогена остатки глюкозо-1-фосфата. Киназа фосфорилазы активируется протеинкиназой А.
Протеинкиназа А
Протеинкиназа А, или цАМФ-зависимая протеинкиназа, относится к семейству ферментов, активность которых зависит от уровня циклического АМФ в клетке. Протеинкиназа А является самой изученной из всех протеинкиназ, её функции разнообразны, она участвует в регуляции метаболизма гликогена, липидов и сахаров, её субстратами могут быть другие протеинкиназы или другие метаболические ферменты.
Протеинкиназа А участвует в цАМФ-стимулируемой транскрипции генов, которые имеют цАМФ-реактивный элемент в регуляторном участке. Повышение концентрации цАМФ ведет к активации протеинкиназы А, которая фосфорилирует транскрипционный фактор CREB по остатку серина 133, CREB связывает своим фосфорилированным участком коактиватор транскрипции и стимулирует транскрипцию.
Молекула протеинкиназы А является холоферментом и в неактивном состоянии является тетрамером — состоит из двух регуляторных и двух каталитических субъединиц. Если уровень цАМФ в клетке низкий, холофермент остается интактным и каталитическая активность отсутствует. Когда концентрация цАМФ в клетке возрастает, цАМФ связывается с двумя сайтами связывания на регуляторных субъединицах, происходят конформационные изменения, диссоциация тетрамера на два каталитически активных димера. Открытые активные центры каталитических субъединиц переносят концевой фосфат молекулы АТФ на остатки серина или треонина.
Протеинкиназы А представлены в многих типах клеток, и проявляют каталитические активности в отношении разных субстратов, поэтому, работа протеинкиназы А и концентрация цАМФ регулируется во многих биохимических путях. Следует отметить, что действие протеинкиназы А, вызванное фосфорилированием, как правило, кратковременное, так как протеинфосфатазы, сопряженные с протеинкиназами, быстро дефосфорилирует мишени.
Гормоны инсулин и глюкагон влияют на работу протеинкиназы А, изменяя уровень цАМФ в клетке по механизму активации рецепторов, связанных с G-белками, через аденилатциклазу. Инсулин активирует аденилатциклазу, повышая концентрацию цАМФ, протеинкиназа А фосфорилирует ферменты ацетил-КоА-карбоксилазу и пируватдегидрогеназу, направляя таким образом ацетил-КоА для синтеза липидов; глюкагон имеет противоположный эффект.
Работа протеинкиназы А регулируется и по механизму отрицательной обратной связи. Одним из субстратов, активируемых протеинкиназой А, является фосфодиэстераза, которая превращает цАМФ в АМФ, таким образом, снижая концентрацию цАМФ и ингибируя протеинкиназу А.
Протеинкиназа B
Геном человека содержит семейство генов Akt1, Akt2, Akt3. Протеинкиназа Akt1 ингибирует процессы апоптоза, принимает участие в регуляции клеточного цикла, индуцирует синтез белка и поэтому является ключевым белком, регулирующим рост тканей, а также отвечает за развитие мышечной гипертрофии. Поскольку продукт гена Akt1 блокирует апоптоз, повышенный уровень экспрессии Akt1 отмечается во многих опухолях. Первоначально Akt1 был охарактеризован как онкоген в трансформирующем ретровирусе AKT8 в 1990 году.
Продукт гена Akt2 является важной сигнальной молекулой в пути передачи сигнала молекулой инсулина, этот белок требуется для транспорта глюкозы.
Показано, что Akt3 преимущественно экспрессируется в мозге. Мыши, лишенные гена Akt3, имеют маленький мозг. Мыши, нокаутные по гену Akt1, но имеющие ген Akt2 имели меньший размер. Так как уровень глюкозы у таких мышей был в норме, была показана роль Akt1 в процессах роста.
Мыши, нокаутные по гену Akt2, но имеющие Akt1, имели задержки роста и фенотипические проявления инсулин-зависимого диабета. Полученные данные указывали на роль Akt2 в проведении сигнала от инсулинового рецептора.
Регуляция активности Akt осуществляется путем связывания фосфолипидов в мембране. Akt содержит PH-домен, который высокоаффинно связывает фосфатидилинозитол-трифосфат или фосфатидилинозитол-дифосфат. PH-домены выполняют функции заякоривания в мембранах. PIP2 может быть фосфорилирован только PIP3-киназами, и только в случае, когда клетка получила сигнал к росту. PIP3-киназы могут быть активированы рецепторами, связанными с G-белками или рецепторами с тирозинкиназной активностью. Только после активации PIP3-киназы фосфорилируют PIP2 в PIP3.
После связывания с PIP3 и закрепления в мембране Akt может быть активирована путем фосфорилирования фосфоинозитол-зависимыми киназами. PDK1 фосфорилирует Akt, mTORC2 стимулирует фосфорилирование PDK1. Активированная Akt далее регулирует фосфорилированием активность многих субстратов. Показано, что Akt может быть активирована и без участия PIP3-киназ.
Соединения, повышающие концентрацию цАМФ могут активировать Akt через протеинкиназу А. Фосфатазы липидов контролируют концентрацию PIP3, например, супрессор опухолей PTEN работает как фосфатаза, и дефосфорилирует PIP3 в PIP2. Akt диссоциирует от плазматической мембраны и активность фермента значительно падает. Протеинфосфатазы контролируют количество фосфорилированного Akt. Фосфатазы PHLPP дефосфорилируют серин 473 в Akt и тем самым инактивируют её.
Akt регулирует многие процессы, направленные на выживание клетки. Например, Akt может фосфорилировать про-апоптотический белок BAD по остатку серина 136, что вызывает диссоциацию BAD из Bcl-2/Bcl-X комплекса и приводит к утере про-апоптотической функции. Также Akt активирует транскрипционный фактор NF-κB, и включает транскрипцию генов выживания.
Akt требуется для инсулин-индуцируемой транслокации транспортера глюкозы 4 в плазматическую мембрану. Киназа-3 гликогенсинтетазы может быть ингибирована фосфорилированием Akt, что вызывает синтез гликогена.
Akt1 также связана с ростом сосудов и развитием опухолей. Недостаточность Akt1 у мышей ингибирует физиологический ангиогенез, но усиливает патологический рост сосудов и опухолей.
Протеинкиназа С
Протеинкиназы С — это семейство протеинкиназ, содержащее порядка десяти изоферментов, которые классифицируют по вторичным посредникам на три семейства: традиционные, или классические, оригинальные, или нестандартные и нетипичные. Традиционным протеинкиназам С для активации требуется Ca, диацилглицерол или фосфатидилхолин.
Оригинальные протеинкиназы С активируются молекулами диацилглицерола, и не требуют Ca. Традиционные и оригинальные протеинкиназы С активируются через сходные пути сигнальной трансдукции, например, с помощью фосфолипазы С. Нетипичные изоформы, не требуют ни Ca, ни диацилглицерола для активации.
Все протеинкиназы С состоят из регуляторного и каталитического доменов, связанных шарнирной областью. Каталитические районы высоко консервативны между разными изоформами, и значительно отличаются от каталитических районов других серин-треониновых протеинкиназ. Консервативность каталитических доменов связано с выполняемыми функциями, различия в регуляторных районах обуславливают различия во вторичных посредниках.
Регуляторный домен на N-конце протеинкиназы С содержит отдельные участки. С1 домен, представленный во всех изоформах протеинкиназы С, имеет сайт связывания диацилглицерола. С2 домен воспринимает ион Ca. Район псевдосвязывания субстрата представляет собой короткую последовательность аминокислот, которые подражают субстрату и занимают участок связывания субстрата в активном центре, делая фермент неактивным.
Ca и диацилглицерол связываются с доменами С2 и С1, соответственно, и вызывают прикрепление протеинкиназы С к плазматической мембране. Взаимодействие с мембраной вызывает освобождение псевдосубстрата из каталитического центра и активирует фермент. Для осуществления подобных аллостерических взаимодействий, протеинкиназе С требуется предварительное фосфорилирование каталитического района.
Протеинкиназа С должна быть предварительно фосфорилирована и для осуществления собственной киназной активности. Молекула протеинкиназы С содержит несколько сайтов фосфорилирования 3-фосфоинозитол-зависимой протеинкиназой-1. После активации протеинкиназа С переносится к плазматической мембране и присоединяются к RACK-белкам, аминокислотная последовательность которых на 47 % гомологична бета-субъединицам G-белков.
Для протеинкиназ С характерен длительный период активности, которая сохраняется, даже если первоначальный сигнал пропал или снизилась концентрация ионов Ca. Это достигается образованием диацилглицерола из фосфатидилхолина с помощью фосфолипазы С.
Последовательность остатков аминокислот в молекуле протеинкиназы С сходна с таковой для протеинкиназы А, и содержит остатки основных аминокислот вблизи остатков серина и треонина, подвергающихся фосфорилированию. Субстратами протеинкиназы С являются следующие белки: MAP-киназы, Raf-киназы, MARCKS. Белки-субстраты протеинкиназы С играют важную роль в поддержании формы клеток, способности к движению, секреции, трансмембранном транспорте, регуляции клеточного цикла. MARCKS вовлечены в процессы экзоцитоза некоторых секреторных пузырьков, содержащих, муцин и хромафин. MARCKS — кислые белки, содержат большое количество остатков аланина, глицина, пролина и глутаминовой кислоты. MARCKS связаны N-концом с липидами мембраны, регулируются ионами Ca, кальмодулином, протеинкиназой С.
VDR — кальцитриоловый рецептор. Рецептор стероидных гормонов из семейства ядерных рецепторов. После активирования молекулой витамина D, образует гетеродимер с ретиноидным-Х-рецептором и связывается с регуляторными элементами на ДНК, изменяя экспрессию генов или снимая репрессоры генов. Глюкокортикоиды снижают экспрессию VDR во всех тканях.
Рецептор эпидермального фактора роста — относится к семейству рецепторов факторов роста, связывающих внеклеточные белковые лиганды и обладает тирозинкиназными активностями. Мутации, затрагивающие EGRF часто могут проявляться в раковом перерождении клетки. После связывания лиганда, рецептор димеризуется, происходит самофосфорилирование по пяти остаткам тирозина на С-конце рецептора, и EGRF приобретает внутриклеточную тирозинкиназную активность.
Последующая активность EGRF связана с инициацией каскада сигнальной трансдукции, активируются MAPK, Akt, JNK — что приводит к синтезу ДНК и пролиферации. Киназный домен также может фосфорилировать другие рецепторы, связанные с EGRF по остаткам тирозина.
Ca/кальмодулин — зависимые протеинкиназы
Са/кальмодулин-зависимые киназы, или СаМ киназы, регулируются Са/кальмодулиновым комплексом. СаМ киназы классифицируют на два класса: специализированные СаМ киназы и многофункциональные СаМ киназы, порядка 2 % белков головного мозга представлены СаМ второго типа.
Кальмодулин — это вездесущий, кальций-связывающий белок, который связывается с многими другими белками и регулирует их активность. Это маленький кислый белок, состоит из 148 аминокислотных остатков, содержит четыре домена связывания кальция.
СаМ служит промежуточным звеном в воспалении, апоптозе, мышечных сокращениях, развитии кратковременной и длительной памяти, росте нервов и иммунном ответе. Кальмодулин экспрессируется во многих типах клеток и находится в цитоплазме, внутри органелл, а также находится в плазматической мембране и мембранах органелл. Многие белки, которые связываются с кальмодулином, не могут сами связывать кальций и используют кальмодулин как «датчик» кальция и компонент системы передачи сигнала.
Кальмодулин также используется для запасания Ca в эндоплазматическом и саркоплазматическом ретикулумах. После связывания кальция молекула кальмодулина претерпевает конформационные изменения, что позволяет молекуле связывать другие белки для осуществления специфического ответа. Молекула кальмодулина может связать до четырёх ионов кальция, может подвергаться посттрансляционной модификации, например, фосфорилированию, ацетилированию, метилированию, протеолизу, причем эти модификации могут модулировать активность СаМ.
Киназа легких цепей миозина. Киназа легких цепей миозина фосфорилирует миозин. Киназа легких цепей миозина имеет ключевое значение в сокращении гладкой мускулатуры. Сокращение гладких мышц может произойти после повышения концентрации кальция в результате притока из саркоплазматического ретикулюма или из внеклеточного пространства. Сперва кальций связывается с кальмодулином, это связывание активирует киназу легких цепей миозина, которая фосфорилирует легкие цепи молекул миозина. Фосфорилирование позволяет молекулам миозина образовывать поперечные мостики и связываться с актиновыми филаментами и стимулирует мышечное сокращение. Данный путь является основным в механизме сокращения гладких мышц, так как гладкие мышцы не содержат тропонинового комплекса, в отличие от поперечно-полосатых.
МАРK
Митоген-активируемые киназы отвечают на внеклеточные стимулы и регулируют многие клеточные процессы. МАРК вовлечены в работу многих неядерных белков — продуктов онкогенов. Внеклеточные стимулы ведут к активации МАРК через сигнальный каскад, который состоит из МАРК, МАРКК и МАРККК. МАР3К активируется внеклеточными стимулами и фосфорилирует МАР2К, затем МАР2К фосфорилированием активирует МАРК. Такой сигнальный МАР-каскад консервативен для эукариот от дрожжей до млекопитающих.
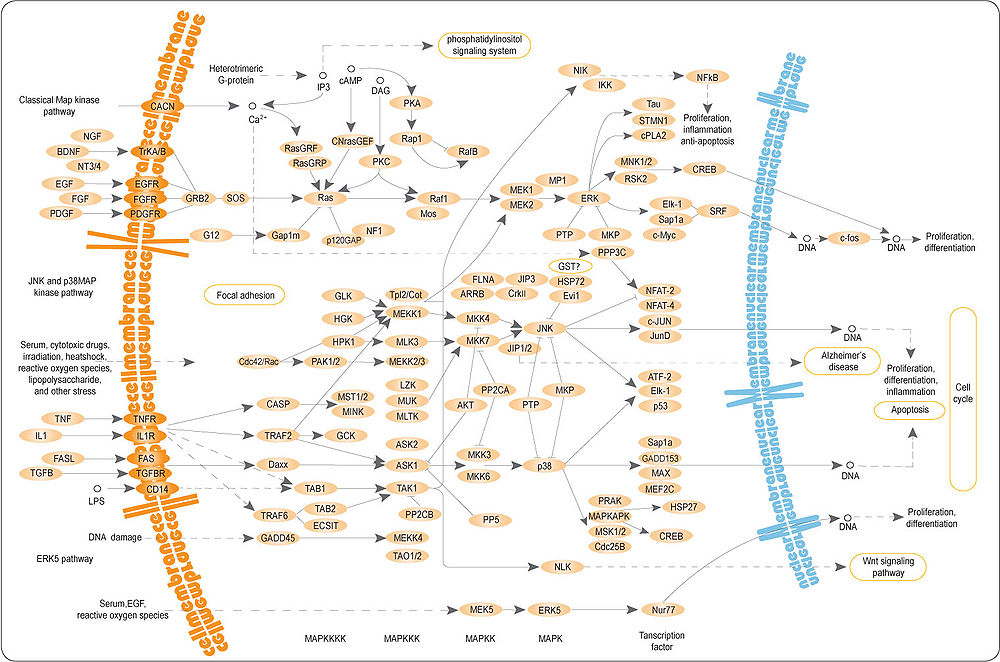
MAPK/ERK-киназы принимают участие в особом пути сигнальной трансдукции. ERK — или классические МАР-киназы, регулируются внеклеточными сигналами.
Рецепторы, связанные с тирозиновыми киназами, активируются внеклеточными лигандами. Связывание EGF с рецептором приводит к фосфорилированию EGFR. Белок GRB2, содержащий SH2-домен, связывается с остатками фосфорилированного тирозина. Белок GRB2 своим SH3-доменом связывается и активирует SOS. Активированный гуанин-нуклеотид заменяющий фактор отщепляет ГДФ от белка Ras, Ras далее может связать ГТФ и активироваться.
Активный Ras активирует RAF-киназу. RAF-киназа фосфорилирует и активирует МЕК, другую серин-треониновую киназу. МЕК фосфорилирует и активирует МАРК. Эта серия киназ от RAF к МЕК и к МАРК является примером каскада протеинкиназ.
Один из эффектов активации МАРК это изменение трансляции мРНК. МАРК фосфорилирует и активирует S6 киназу 40S рибосомных белков. RSK фосфорилирует рибосомный белок S6, и вызывает его диссоциацию от рибосомы.
МАРК регулирует активности нескольких транскрипционных факторов, например, C-myc. МАРК регулирует активность генов, контролирующих клеточный цикл.
Просмотров: 16138
|
|