Биология - Тилакоид - Функции тилакоидов
09 февраля 2011Оглавление:
1. Тилакоид
2. Образование тилакоидов
3. Выделение и фракционирование тилакоидов
4. Функции тилакоидов
5. Мембраны тилакоидов цианобактерий
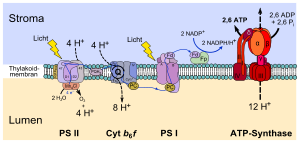
В тилакоидах осуществляются следу светозависимые реакции фотосинтеза:
- Светозависимое расщепление воды, в результате которого происходит синтез молекул кислорода;
- Перенос протонов через тилакоидную мембрану, связанный с электронтранспортной цепью фотосистем и цитохромного комплекса b6f;
- Синтез АТФ, выполняемый АТФ-синтазой с использованием протонного градиента.
Фотолиз воды
Первый этап фотосинтеза — это расщепление воды под воздействием света. Эта реакция поставляет электроны для фотосинтетических электронтранспортных цепей, а также протоны для создания протонного градиента. Реакция расщепления воды происходит на стороне тилакоидной мембраны, обращённой к люмену, и происходит с затратами энергии, полученной фотосистемами от солнечного света. Интересно отметить, что это окисление воды происходит с выделением O2 как побочного продукта, который сбрасывается в атмосферу и затем может быть использован другими организмами для дыхания.
Электронтранспортная цепь
В процессе фотосинтеза использованы две разновидности транспорта электроннов:
- Нециклический электронный транспорт или нециклическое фотофосфорилирование, при котором производится NADPH, протоны H и АТФ;
- Циклический электронный транспорт или циклическое фотофосфорилирование, при котором производится только АТФ.
Нециклическая разновидность транспорта задействует обе фотосистемы, тогда как циклическая происходит только с использованием фотосистемы I.
- Фотосистема I использует энергию света для восстановления NADP до NADPH + H. Она активна в обеих разновидностях электронного транспорта. В циклическом режиме возбужденный электрон отправляется по цепочке, которая в конце возвращает его обратно к хлорофиллу, который сообщил ему энергию возбуждения.
- Фотосистема II использует энергию света для расщепления молекул воды с выходом электронов, протонов и молекулярного кислорода. Она задействована только в нециклическом транспорте. Электроны в добываются этой системой из окисленного 2H2O и удаляются из нее с NADP, когда он восстанавливается до NADPH.
Электрохимический потенциал
Основанной функцией тилакоидной мембраны и ее интегральных фотосистем является создание электрохимического потенциала. Переносчики электронов, участвующие в электронном транспорте, используют некоторую часть энергии электронов для перекачки протонов из стромы в просвет тилакоида. Во время фотосинтеза вещество люмена приобретает кислую реакцию вплоть до pH 4. Это соответствует 10 000-кратному градиенту концентрации протонов поперек тилакоидной мембраны.
Источник протонного градиента
Протоны в просвет поступают из трех источников:
- Фотолиз воды, осуществляемый фотосистемой II, в процессе которого вода в люмене тилакоида окисляется с выходом молекулярного кислорода, протонов и электронов.
- Передача электрона от фотосистемы II к пластохинону во время светозависимой реакции фотосинтеза, входящей в нециклическую электронтранспортную цепь, потребляет два протона из стромы. Они переносятся в просвет тилакоида, когда восстановленный пластохинон окисляется цитохромным комплексом b6f на стороне тилакоидной мембраны, обращенной к просвету. От пластохинона электроны передаются цитохромному комплексу b6f, который напоминает цитохром bc1.
- Восстановление пластохинона ферредоксином в процессе циклического электронного транспорта также вызывает перенос двух протонов из стромы в просвет тилакоида.
Протонный градиент также поддерживается потреблением протонов в строме при восстановлении NADP до NADPH, осуществляемом NADP-редуктазой.
Синтез АТФ
Молекулярный механизм синтеза АТФ в хлоропластах похож на аналогичный механизм в митохондриях. Он получает необходимую энергию от протон-движущей силы. Однако хлоропласты используют большей частью на электрохимический потенциал ПДС. ПДС слагается из химического потенциала протонов и трансмембранного электрического потенциала.
По сравнению с внутренними мембранами митохондрий, которые обладают существенно более высоким мембранным потенциалом, обусловленным разделением зарядов, градиент заряда на тилакоидных мембранах невелик. В то же время это компенсируется 10000-кратным градиентом концентрации протонов, который гораздо выше, нежели 10-кратный у митохондрий. Общий электрохимический потенциал между просветом тилакоида и стромой достаточно велик, чтобы подпитывать работу АТФ-синтазы. Когда протоны выходят обратно в строму в область сниженной концентрации через канал в АТФ-синтазе, происходит реакция синтеза АТФ. Именно через протонный градиент светозависимые реакции соединены с синтезом АТФ.
Просмотров: 23389
|
|