Биология - Бактерии - Строение
09 февраля 2011Оглавление:
1. Бактерии
2. Термин
3. Строение
4. Способы передвижения и раздражимость
5. Метаболизм
6. Размножение и устройство генетического аппарата
7. Клеточная дифференциация
8. Классификация
9. Происхождение, эволюция, место в развитии жизни на Земле
10. Экология
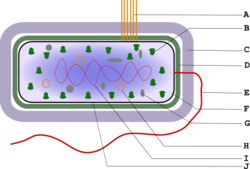
Подавляющее большинство бактерий одноклеточны. По форме клеток они могут быть округлыми, палочковидными, извитыми, реже — звёздчатыми, тетраэдрическими, кубическими, C- или O-образными. Формой определяются такие способности бактерий, как прикрепление к поверхности, подвижность, поглощение питательных веществ. Отмечено, например, что олиготрофы, то есть бактерии, живущие при низком содержании питательных веществ в среде, стремятся увеличить отношение поверхности к объёму, например, с помощью образования выростов.
Из обязательных клеточных структур выделяют три:
- нуклеоид
- рибосомы
- цитоплазматическая мембрана
С внешней стороны от ЦПМ находятся несколько слоёв, называемых клеточной оболочкой, а также поверхностные структуры. ЦПМ и цитоплазму объединяют вместе в понятие протопласт.
Строение протопласта
ЦПМ ограничивает содержимое клетки от внешней среды. Гомогенная фракция цитоплазмы, содержащая набор растворимых РНК, белков, продуктов и субстратов метаболических реакций, названа цитозолем. Другая часть цитоплазмы представлена различными структурными элементами.
Одним из основных отличий клетки бактерий от клетки эукариот является отсутствие ядерной мембраны и, строго говоря, отсутствие вообще внутрицитоплазматических мембран, не являющихся производными ЦПМ. Однако у разных групп прокариот имеются локальные впячивания ЦПМ — мезосомы, выполняющие в клетке разнообразные функции и разделяющие её на функционально различные части. У многих фотосинтезирующих бактерий существует развитая сеть производных от ЦПМ фотосинтетических мембран. У пурпурных бактерий они сохранили связь с ЦПМ, легко обнаруживаемую на срезах под электронным микроскопом, у цианобактерий эта связь либо трудно обнаруживается, либо утрачена в процессе эволюции. В зависимости от условий и возраста культуры фотосинтетические мембраны образуют различные структуры — везикулы, хроматофоры, тилакоиды.
Вся необходимая для жизнедеятельности бактерий генетическая информация содержится в одной ДНК, чаще всего имеющей форму ковалентно замкнутого кольца. Она в одной точке прикреплена к ЦПМ и помещается в структуре, обособленной, но не отделённой мембраной от цитоплазмы, и называемой нуклеоид. ДНК в развёрнутом состоянии имеет длину более 1 мм. Бактериальная хромосома представлена обычно в единственном экземпляре, то есть практически все прокариоты гаплоидны, хотя в определённых условиях одна клетка может содержать несколько копий своей хромосомы, а Burkholderia cepacia имеет три разных кольцевых хромосомы. Рибосомы прокариот также отличны от таковых у эукариот и имеют константу седиментации 70 S.
Помимо этих структур, в цитоплазме также могут находиться включения запасных веществ.
Клеточная оболочка и поверхностные структуры
Клеточная стенка — важный структурный элемент бактериальной клетки, однако необязательный. Искусственным путём были получены формы с частично или полностью отсутствующей клеточной стенкой, которые могли существовать в благоприятных условиях, однако иногда утрачивали способность к делению. Известна также группа природных не содержащих клеточной стенки бактерий — микоплазм.
У бактерий существует два основных типа строения клеточной стенки, свойственных грамположительным и грамотрицательным видам.
Клеточная стенка грамположительных бактерий представляет собой гомогенный слой толщиной 20—80 нм, построенный в основном из пептидогликана с меньшим количеством тейхоевых кислот и небольшим количеством полисахаридов, белков и липидов. В клеточной стенке имеются поры диаметром 1—6 нм, которые делают её проницаемой для ряда молекул.
У грамотрицательных бактерий пептидогликановый слой неплотно прилегает к ЦПМ и имеет толщину лишь 2—3 нм. Он окружён наружной мембраной, имеющей, как правило, неровную, искривлённую форму. Между ЦПМ, слоем пептидогликана и внешней мембраной имеется пространство, называемое периплазматическим, и заполненное раствором, включающим в себя транспортные белки и ферменты.
С внешней стороны от клеточной стенки может находиться капсула — аморфный слой, сохраняющий связь со стенкой. Слизистые слои не имеют связи с клеткой и легко отделяются, чехлы же не аморфны, а имеют тонкую структуру. Однако между этими тремя идеализированными случаями есть множество переходных форм.
Бактериальных жгутиков может быть от 0 до 1000. Возможны как варианты расположения одного жгутика у одного полюса, пучка жгутиков у одного или двух полюсов, так и многочисленные жгутики по всей поверхности клетки. Толщина жгутика составляет 10—20 нм, длина — 3—15 мкм. Его вращение осуществляется против часовой стрелки с частотой 40—60 об/с.
Помимо жгутиков, среди поверхностных структур бактерий необходимо назвать ворсинки. Они тоньше жгутиков и необходимы для прикрепления бактерии к субстрату, принимают участие в транспорте метаболитов, а особые ворсинки — F-пили, нитевидные образования, более тонкие и короткие, чем жгутики, — необходимы клетке-донору для передачи реципиенту ДНК при конъюгации.
Размеры
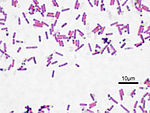
Размеры бактерий в среднем составляют 0,5—5 мкм. Escherichia coli, например, имеет размеры 0,3—1 на 1—6 мкм, Staphylococcus aureus — диаметр 0,5—1 мкм, Bacillus subtilis — 0,75 на 2—3 мкм. Крупнейшей из известных бактерий является Thiomargarita namibiensis, достигающая размера в 750 мкм. Второй является Epulopiscium fishelsoni, имеющая диаметр 80 мкм и длину до 700 мкм и обитающая в пищеварительном тракте хирурговой рыбы Acanthurus nigrofuscus. Achromatium oxaliferum достигает размеров 33 на 100 мкм, Beggiatoa alba — 10 на 50 мкм. Спирохеты могут вырастать в длину до 250 мкм при толщине 0,7 мкм. В то же время к бактериям относятся самые мелкие из имеющих клеточное строение организмов. Mycoplasma mycoides имеет размеры 0,1—0,25 мкм, что соответствует размеру крупных вирусов, например, табачной мозаики, коровьей оспы или гриппа. По теоретическим подсчётам, сферическая клетка диаметром менее 0,15—0,20 мкм становится неспособной к самостоятельному воспроизведению, поскольку в ней физически не могут поместиться все необходимые биополимеры и структуры в достаточном количестве.
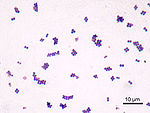
Однако были описаны нанобактерии, имеющие размеры меньше «допустимых» и сильно отличающиеся от обычных бактерий. Они, в отличие от вирусов, способны к самостоятельному росту и размножению. Они пока мало изучены, и их живая природа ставится под сомнение.
При линейном увеличении радиуса клетки её поверхность возрастает пропорционально квадрату радиуса, а объём — пропорционально кубу, поэтому у мелких организмов отношение поверхности к объёму выше, чем у более крупных, что означает для первых более активный обмен веществ с окружающей средой. Метаболическая активность, измеренная по разным показателям, на единицу биомассы у мелких форм выше, чем у крупных. Поэтому небольшие даже для микроорганизмов размеры дают бактериям и археям преимущества в скорости роста и размножения по сравнению с более сложноорганизованными эукариотами и определяют их важную экологическую роль.
Многоклеточность у бактерий
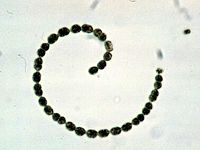
Одноклеточные формы способны осуществлять все функции, присущие организму, независимо от соседних клеток. Многие одноклеточные прокариоты склонны к образованию клеточных агрегатов, часто скреплённых выделяемой ими слизью, эти агрегаты получили название биоплёнки. Чаще всего это лишь случайное объединение отдельных организмов, но в ряде случаев временное объединение связано с осуществлением определённой функции, например, формирование плодовых тел миксобактериями делает возможным развитие цист, при том что единичные клетки не способны их образовывать. Подобные явления наряду с образованием одноклеточными эубактериями морфологически и функционально дифференцированных клеток — необходимые предпосылки для возникновения у них истинной многоклеточности.
Многоклеточный организм должен отвечать следующим условиям:
- его клетки должны быть агрегированы,
- между клетками должно осуществляться разделение функций,
- между агрегированными клетками должны устанавливаться устойчивые специфические контакты.
Многоклеточность у прокариот известна, наиболее высокоорганизованные многоклеточные организмы принадлежат к группам цианобактерий и актиномицетов. У нитчатых цианобактерий описаны структуры в клеточной стенке, обеспечивающие контакт двух соседних клеток — микроплазмодесмы. Показана возможность обмена между клетками веществом и энергией. Некоторые из нитчатых цианобактерий содержат помимо обычных вегетативных клеток функционально дифференцированные: акинеты и гетероцисты. Последние осуществляют фиксацию азота и интенсивно обмениваются метаболитами с вегетативными клетками.
Просмотров: 43521
|
|